Koder innenfor koder: Hvordan dobbelt-bruk kodoner utfordrer statistiske metoder for å slutte til naturlig utvalg
Oversatt herfra:
 Den doble funksjonen til den genetiske koden peker mot design
Den doble funksjonen til den genetiske koden peker mot design
1. Det er funnet et 'overlappende språk' i den genetiske koden
2. Det ene språket beskriver hvordan proteiner lages, og det andre hjelper med å styre genetisk aktivitet i celler. Det ene språket er skrevet oppå det andre, og derfor forble dette andre språket uoppdaget så lenge.
3. Originalpapiret av Stergachis et al. skriver om 'evolusjonære begrensninger' av de overlappende kodene. De skrev: "Våre resultater indikerer at samtidig koding av aminosyre og regulatorisk informasjon i eksoner er et viktig funksjonelt trekk ved komplekse genomer. Informasjonsarkitekturen til den mottatte genetiske koden er optimalisert for plassering av tilleggsinformasjon over seg selv. TF [TranskripsjonsFaktor] binding i eksoner kan tjene flere funksjonelle roller.
4. I følge forskningen begrenser eller eliminerer naturlig utvalg endring (rensende utvalg) og er ikke nyttig for å skape nye organer eller funksjoner.
5. Ordene: informasjon, arkitektur, optimalisert og funksjon refererer alltid, og bare, til en intelligent agent med tenkevne følelse og vilje. Andre foreslåtte agenter kan ikke på egen hånd gi informasjon, designe, optimalisere eller utføre oppgaver. Dette har aldri vist seg å være mulig.
7. En slik intelligent designet kompleks genetisk kode med doble eller til og med tredobbelte funksjoner kunne bare blitt opprettet av en strålende superintelligent designer.
Foruten at det er overlappende koding som er hierarkisk over kodingen av DNA, finner vi at det også er overlappende koding innen DNA:
Flere genetiske koder som overlapper hverandre, reduserer sterkt sannsynligheten for gunstig mutasjon (mai 2013)
Utdrag: I det siste tiåret har vi oppdaget enda et aspekt av det flerdimensjonale genomet. Vi vet nå at DNA-sekvenser vanligvis er 'flerfunksjonelle' [38]. Trifanov hadde tidligere beskrevet minst 12 genetiske koder som et gitt nukleotid kan bidra til [39,40], og viste at et gitt basepar kan bidra til flere overlappende koder samtidig. Det første beviset på overlappende proteinkodende sekvenser i virus forårsaket ikke så lite oppstyr, men siden har det blitt anerkjent som typisk. I følge Kapronov et al., "Er det ikke uvanlig at et enkelt basepar kan være en del av et intrikat nettverk av flere isoformer med overlappende sans og antisense-transkripsjoner, hvorav flertallet ikke er kommentert" [41]. ENCODE-prosjektet [42] har bekreftet at dette fenomenet er allestedsnærværende i høyere genomer, hvor en gitt DNA-sekvens rutinemessig koder for flere overlappende meldinger, noe som betyr at et enkelt nukleotid kan bidra til to eller flere genetiske koder. Senest har Itzkovitz et al. analyserte proteinkodende regioner på 700 arter og viste at så å si alle former for liv har omfattende overlappende informasjon i genomene [43].
Lenke.
 "Det er rikelig med bevis for at de fleste DNA-sekvenser er flerfunksjonelle, og derfor er flerbegrensede. Dette faktum har blitt demonstrert grundig av Trifonov (1989). For eksempel koder de fleste menneskelige kodende sekvenser for to forskjellige RNAer, lest i motsatte retninger, dvs. begge DNA-strengene er transkribert (Yelin et al., 2003). Noen sekvenser koder for forskjellige proteiner, avhengig av hvor oversettelsen startes og hvor leserammen begynner (dvs. gjennomleste proteiner). Noen sekvenser koder for forskjellige proteiner basert på alternativ mRNA-spleising. Noen sekvenser tjener samtidig for proteinkoding og fungerer også som interne transkripsjonspromotorer. Noen sekvenser koder for både en proteinkodende og en proteinbindende region. Alu-elementer og replikasjonsopprinnelse finnes i funksjonelle promotorer og i eksoner. I utgangspunktet er alle DNA-sekvenser begrenset av isochore-krav (regionalt GC-innhold), 'ord' -innhold (artsspesifikke profiler av di-, tri- og tetra-nukleotidfrekvenser) og nukleosombindingssteder (dvs. Alt DNA må trekkes sammen) . Selektiv sammentrekking er tydelig involvert i genregulering, og selektiv nukleosom-binding styres av spesifikke DNA-sekvensmønstre - som må gjennomsyre hele genomet. Til slutt, gjør sannsynligvis alle sekvenser det de gjør, selv om de også påvirker den generelle avstanden og DNA-folding/arkitekturen - som helt klart er sekvensavhengig. For å forklare den utrolige mengden informasjon som på en eller annen måte må pakkes inn i genomet (gitt livets ekstreme kompleksitet), må vi virkelig anta at det er enda høyere nivåer av organisasjon og informasjon kryptert i genomet. For eksempel er det et helt annet nivå av organisering på epigenetisk nivå (Gibbs 2003). Det ser også ut til å være en omfattende sekvensavhengig tredimensjonal organisasjon innen kromosomer og hele kjernen (Manuelides, 1990; Gardiner, 1995; Flam, 1994). Trifonov (1989), har vist at alle DNA-sekvenser i genomet sannsynligvis krypterer flere 'koder' (opptil 12 koder)."
"Det er rikelig med bevis for at de fleste DNA-sekvenser er flerfunksjonelle, og derfor er flerbegrensede. Dette faktum har blitt demonstrert grundig av Trifonov (1989). For eksempel koder de fleste menneskelige kodende sekvenser for to forskjellige RNAer, lest i motsatte retninger, dvs. begge DNA-strengene er transkribert (Yelin et al., 2003). Noen sekvenser koder for forskjellige proteiner, avhengig av hvor oversettelsen startes og hvor leserammen begynner (dvs. gjennomleste proteiner). Noen sekvenser koder for forskjellige proteiner basert på alternativ mRNA-spleising. Noen sekvenser tjener samtidig for proteinkoding og fungerer også som interne transkripsjonspromotorer. Noen sekvenser koder for både en proteinkodende og en proteinbindende region. Alu-elementer og replikasjonsopprinnelse finnes i funksjonelle promotorer og i eksoner. I utgangspunktet er alle DNA-sekvenser begrenset av isochore-krav (regionalt GC-innhold), 'ord' -innhold (artsspesifikke profiler av di-, tri- og tetra-nukleotidfrekvenser) og nukleosombindingssteder (dvs. Alt DNA må trekkes sammen) . Selektiv sammentrekking er tydelig involvert i genregulering, og selektiv nukleosom-binding styres av spesifikke DNA-sekvensmønstre - som må gjennomsyre hele genomet. Til slutt, gjør sannsynligvis alle sekvenser det de gjør, selv om de også påvirker den generelle avstanden og DNA-folding/arkitekturen - som helt klart er sekvensavhengig. For å forklare den utrolige mengden informasjon som på en eller annen måte må pakkes inn i genomet (gitt livets ekstreme kompleksitet), må vi virkelig anta at det er enda høyere nivåer av organisasjon og informasjon kryptert i genomet. For eksempel er det et helt annet nivå av organisering på epigenetisk nivå (Gibbs 2003). Det ser også ut til å være en omfattende sekvensavhengig tredimensjonal organisasjon innen kromosomer og hele kjernen (Manuelides, 1990; Gardiner, 1995; Flam, 1994). Trifonov (1989), har vist at alle DNA-sekvenser i genomet sannsynligvis krypterer flere 'koder' (opptil 12 koder)."
Dr. John Sanford; Genetisk entropi 2005
Videre er det veldig gode matematiske grunner til at overlappende koding i DNA vil forhindre at en skapning noen gang blir forandret til en annen skapning.
Flere overlappende genetiske koder reduserer sannsynligheten for gunstig mutasjon sterkt John C. Sanford 4. mai 2013
Konklusjoner: Analysen vår bekrefter matematisk hva som virker intuitivt åpenbart - flere overlappende koder i genomet må radikalt endre våre forventninger til frekvensen av gunstige mutasjoner. Når antallet overlappende koder øker, synker frekvensen av potensiell gunstig mutasjon eksponentielt og nærmer seg null. Derfor indikerer det nye beviset for allestedsnærværende overlappende koder i høyere genomer sterkt at gunstige mutasjoner skal være ekstremt sjeldne. Dette beviset kombinert med økende bevis for at biologiske systemer er høyst optimalisert, og bevis for at bare gunstige mutasjoner med relativt stor innvirkning kan forsterkes effektivt ved naturlig seleksjon, får oss til å konkludere med at mutasjoner som både er valgbare og utvetydig fordelaktige, må være forsvinnende sjeldne. Denne konklusjonen reiser alvorlige spørsmål. Hvordan kan slike forsvinnende sjeldne fordelaktige mutasjoner noen gang være tilstrekkelig for genom-bygging? Hvordan kan genetisk degenerasjon noen gang avverges, gitt den kontinuerlige akkumuleringen av skadelige mutasjoner med lite innvirkning?
http://www.worldscientific.com/doi/pdf/10.1142/9789814508728_0006
Bilde: Eks. på palindromer
 En veldig enkel måte å forstå den monumentale murveggen ethvert evolusjonært scenario står overfor med flere overlappende kodinger som finnes i DNA, er med følgende puslespill funnet på side 141 i boka 'Genetic Entropy' av Dr. Sanford.
En veldig enkel måte å forstå den monumentale murveggen ethvert evolusjonært scenario står overfor med flere overlappende kodinger som finnes i DNA, er med følgende puslespill funnet på side 141 i boka 'Genetic Entropy' av Dr. Sanford.
S A T O R
A R E P O
T E N E T
O P E R A
R O T A S
Som er oversatt;
THE SOWER NAMED AREPO HOLDS THE WORKING OF THE WHEELS.
Dette eldgamle puslespillet, som går tilbake til minst 79 e.Kr., blir det samme, lest fire forskjellige veier, men vi vil konsekvent ødelegge de andre 3 avlesningene av meldingen med den nye mutasjonen (unntatt for den midtre).
Dette er hva som menes når det sies at et polyfunksjonelt genom er poly-begrenset for eventuelle tilfeldige mutasjoner.
Dette flerbegrensede prinsippet er grunnen til at vi aldri ser den ubegrensede plastisiteten i organismer som var og er forestilt av Darwin og hans tilhengere, og er også grunnen til at tilfeldige mutasjoner, som har effekter som er store nok til at vi er i stand til å måle dem, nesten alltid er skadelige i effekten som måles:
"Uansett hva vi prøver å gjøre innenfor en gitt art, når vi snart grenser som vi ikke kan gjennombrudd. Det finnes en vegg på hver side av hver art. Den veggen er DNA-kodingen, som tillater stort utvalg i den (innenfor genbassenget eller genotypen til en art) - men ingen utgang gjennom den veggen. Darwins gradualisme er avgrenset av interne begrensninger, utover hvilke seleksjon er ubrukelig."
R. Milner, Encyclopedia of Evolution (1990)
 Videre, på morfologisk og atferdsmessig nivå, finner vi at sjimpanser og mennesker er langt mer forskjellige enn det man vanligvis tror.
Videre, på morfologisk og atferdsmessig nivå, finner vi at sjimpanser og mennesker er langt mer forskjellige enn det man vanligvis tror.
Faktisk sa King og Wilson, som var de første som antydet at vi er 98% like sjimpanser på genetisk nivå, at siden den morfologiske og atferdsmessige forskjellen mellom sjimpanser og mennesker er så stor, er den morfologiske og atferdsmessige forskjellen mellom mennesker og aper må skyldes variasjoner i deres genomiske reguleringssystemer, siden en slik likhet i proteinkodende regioner åpenbart ikke kunne forklare den store morfologiske og atferdsmessige forskjellen mellom sjimpanser og mennesker.
I 'Science', 1975, var M-C King og A.C. Wilson de første til å publisere en artikkel som estimerte graden av likhet mellom det menneskelige og sjimpanse-genet. Dette dokumenterte graden av genetisk likhet mellom de to! Studien, ved hjelp av et begrenset datasett, fant at vi var langt mer like enn man antok var mulig den gangen. Derfor må vi være ens med aper, ikke sant? Men ... i den andre delen av artikkelen beskriver King og Wilson ærlig manglene ved et slikt resonnement:
"Den molekylære likheten mellom sjimpanser og mennesker er ekstraordinær fordi de skiller seg mye mer ut enn søsken i anatomi og livsstil. Selv om mennesker og sjimpanser er ganske like i strukturen til brystkassen og armene, skiller de seg vesentlig ikke bare i hjernestørrelse, men også i bekken, fot og kjever, samt i relative lengder på lemmer og tær" (38).
Mennesker og sjimpanser er også forskjellige i mange andre anatomiske henseender, i den grad at nesten alle bein i sjimpansens kropp lett kan skilles i form eller størrelse fra dets menneskelige motstykke (38).
I forbindelse med disse anatomiske forskjellene er det selvfølgelig store forskjeller i holdning , bevegelsesmåte, måte å skaffe mat på og kommunikasjons-måter. På grunn av disse store forskjellene i anatomi og livsstil plasserer biologer de to artene ikke bare i separate slekter, men i separate familier (39). Så det ser ut til at molekylære og organisme-metoder for å evaluere forskjellen på den sjimpanse menneskelige gir ganske forskjellige konklusjoner (40). "
King og Wilson fortsatte med å antyde at det morfologiske og atferdsmessige mellom mennesker og aper må skyldes variasjoner i deres genomiske reguleringssystemer.
David Berlinski - Djevelens villfarelse - Side 162 & 163
'Evolution at Two Levels in Humans and Chimpanzees' Mary-Claire King; A. C. Wilson - 1975
Lenke.
Faktisk så store er de anatomiske forskjellene mellom mennesker og sjimpanser at en darwinist, siden griser er anatomisk nærmere mennesker enn sjimpanser, faktisk foreslått at en sjimpanse og gris parret med hverandre, og det er det som til slutt ga opphav til mennesker. (Jeg antar at selv hybridisering ikke kjenner til grenser, i hodet til noen darwinister).
Videre publiserte Physorg en påfølgende artikkel som viser at grisechimpanse-hybridteorien for menneskelig opprinnelse er mye vanskeligere å skyte ned enn noen andre darwinister, som motsatte seg McCarthys radikale teori, hadde først antatt at det ville være: Menneskelige hybrider: en nærmere titt på teorien og bevisene - 25. juli 2013
Utdrag: Det var betydelig nedfall, både positivt og negativt, fra vår første historie som dekker den radikale grisechimpanse-hybridteorien som ble fremsatt av Dr. Eugene McCarthy ... i det store og hele hadde de som kom imot teorien overraskende lite vitenskap å tilby i deres noen ganger personlige angrep mot McCarthy.
.... Under den alternative hypotesen (mennesker er ikke grise-sjimpanse-hybrider), er antagelsen at mennesker og sjimpanser er like fjernt fra griser. Du forventer derfor at 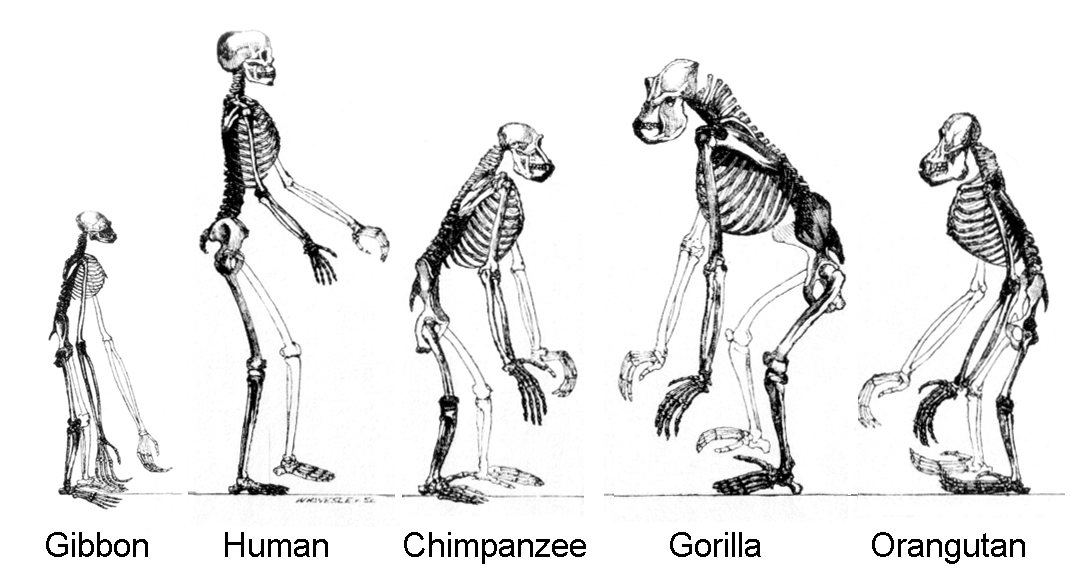 sjimpanseegenskaper som ikke er sett hos mennesker, vil være til stede i griser i omtrent samme hastighet som menneskelige egenskaper som ikke finnes i sjimpanser. Da han søkte i litteraturen etter trekk som skiller mennesker og sjimpanser og samlet en lang liste over slike trekk, fant han imidlertid at det alltid var mennesker som lignet griser med hensyn til disse egenskapene. Dette funnet er uforenlig med muligheten for at mennesker ikke er svin-sjimpansehybrider, det vil si at det avviser hypotesen. .. http://phys.org/news/2013-07-human-hybrids-closer-theory-evidence.html
sjimpanseegenskaper som ikke er sett hos mennesker, vil være til stede i griser i omtrent samme hastighet som menneskelige egenskaper som ikke finnes i sjimpanser. Da han søkte i litteraturen etter trekk som skiller mennesker og sjimpanser og samlet en lang liste over slike trekk, fant han imidlertid at det alltid var mennesker som lignet griser med hensyn til disse egenskapene. Dette funnet er uforenlig med muligheten for at mennesker ikke er svin-sjimpansehybrider, det vil si at det avviser hypotesen. .. http://phys.org/news/2013-07-human-hybrids-closer-theory-evidence.html
Relevant stoff om dette -her.
Selvfølgelig er det ikke ett eneste fnugg av empirisk bevis som antyder at slike radikalt forskjellige skapninger, som griser og sjimpanser, noen gang med hell kunne produsere levedyktige avkom.
Men akk, når teorien din i utgangspunktet er bygget på historiefortelling, (og ikke på noen reelle empiriske bevis), så kommer du selvfølgelig ikke til å kunne skyte ned en annen det bare er slik historie' bare fordi du ikke liker hvordan fortellingen motsier din foretrukne fortelling at mennesket stammer fra aper:
"Vi har alle sett den kanoniske paraden av aper, hver og en blir mer menneskelig. Vi vet at, som en skildring av evolusjon, er denne oppstillingen tosh (dvs. tull). Likevel holder vi oss til det. Ideer om hvordan menneskelig evolusjon burde ha vært, farger fremdeles debatten vår."
Henry Gee, redaktør av Nature (478, 6. oktober 2011, side 34, doi: 10.1038 / 478034a)
I en videre merknad til King og Wilsons observasjon, at 'nesten alle bein i sjimpansens kropp er lett å skille i form eller størrelse fra sitt menneskelige motstykke', plasserer denne observasjonen av King og Wilson i seg selv en annen alvorlig restriksjon for den darwinistiske evolusjonen. Det setter nok en gang spørsmålstegn ved hele teorien.
Enkelt sagt, siden nesten alle bein lett kan skelnes mellom sjimpanser og mennesker, er det flere samtidige koordinerte endringer som kreves i stedet for bare individuelle endringer, som det er tenkt i darwinismen, så for å forhindre katastrofale resultater:
Forbinde punktene: Modellering av funksjonell integrasjon i biologiske systemer - 11. juni 2010
Utdrag: "Hvis en ingeniør endrer lengden på stempelstengene i en forbrenningsmotor, men ikke endrer veivakselen tilsvarende, vil ikke motoren starte. Tilsvarende er utviklingsprosesser så tett integrert tidsmessig og romlig at en endring tidlig i utviklingen vil kreve en rekke andre koordinerte endringer i separate, men funksjonelt sammenhengende utviklingsprosesser nedstrøms "(1)
Lenke.
"Dette er problemet jeg har med neo-darwinister: De lærer at det som skaper noe nytt er akkumulering av tilfeldige mutasjoner i DNA, i en retning satt av naturlig seleksjon. Hvis du vil ha større egg, fortsetter du å velge høner som legger de største eggene, og du får større og større egg. Men du får også høner med defekte fjær og vaklende ben. Naturlig utvalg eliminerer og opprettholder kanskje, men det skaper ikke….
(Sitert i "Discover Interview: Lynn Margulis sier at hun ikke er kontroversiell, hun har rett", Discover Magazine, s. 68 (april, 2011).)
"Det reelle antall variasjoner er færre enn forventet ,,. Det er ingen blåøyede Drosophila, ingen fugler eller skilpadder som føder levende unger , ingen pattedyr med seks ben osv. Slike observasjoner provoserer evolusjonære begreper som ikke er darwinistiske. Darwin prøvde ganske uten hell å løse problemet med motsetningene mellom modellen for tilfeldig variabilitet og eksistensen av begrensninger. Han prøvde å skjule denne komplikasjonen ved å sitere mange fakta om andre fenomener. Forfatterne av de moderne versjonene av darwinismen fulgte denne strategien, slik at problemet kunne vedvare. ... Han ble imidlertid tvunget til å innrømme noen tilfeller der det var umulig å skape noe mennesker kunne ønske seg. Da de engelske bøndene for eksempel bestemte seg for å skaffe kuer med tykk skinke, forlot de snart dette forsøket siden de omkom for ofte under fødselen. Slike tilfeller provoserte tydeligvis en idé om begrensningene til variabilitet ... [Hvis du har tid, les hele den følgende artikkelen, som konkluderer] Problemet med begrensningene i variasjon ble ikke løst verken innenfor rammen av riktig Darwinistisk teori, eller innenfor rammen av moderne darwinisme. " (IGOR POPOV, THE PROBLEM OF CONSTRAINTS ON VARIATION, FROM DARWIN TO THE PRESENT, 2009.
Bilde: Karakteristiska ved design-argumentet
 Kanskje det er grunnen til at så mange ingeniører støtter intelligent design, siden de lett kan se umuligheten av 'engineering problem' for darwinistiske prosesser. Design må nemlig implementeres ovenfra og ned, med alle delene koordinert med hverandre, for å unngå katastrofale resultater for systemet som helhet.
Kanskje det er grunnen til at så mange ingeniører støtter intelligent design, siden de lett kan se umuligheten av 'engineering problem' for darwinistiske prosesser. Design må nemlig implementeres ovenfra og ned, med alle delene koordinert med hverandre, for å unngå katastrofale resultater for systemet som helhet.
Videre, i ytterligere merknad til King og Wilsons påstand om at den morfologiske og atferdsmessige forskjellen mellom mennesker og aper må skyldes variasjoner i deres genomiske reguleringssystemer, (siden den genetiske likheten åpenbart ikke kan forklare den store morfologiske og atferdsmessige forskjellen mellom sjimpanser og mennesker) , finner vi at det faktisk er i de genetiske reguleringsregionene vi finner 'størrelsesordener' og 'artsspesifikke' forskjeller mellom ikke bare sjimpanser og mennesker, men også i andre arter: Bare en påminnelse om at genetisk likhet er langt mer utbredt, på tvers av veldig forskjellige arter, enn darwinister forventet at den genetiske likheten skulle være
Hai og menneskelige proteiner er 'utrolig like': hai er nærmere menneske enn sebrafisk - 9. desember 2013
Utdrag: "Vi var veldig overrasket over å finne at haier i mange kategorier proteiner har flere likheter med mennesker enn sebrafisk," sa Stanhope. "Selv om haier og benfisk ikke er nært beslektede, er de likevel begge fiskene .. mens pattedyr har veldig forskjellige anatomier og fysiologier.
Kenguruegener nær menneskers
Utdrag: Australias kenguruer ligner genetisk på mennesker .... "Det er noen forskjeller, vi har noen flere av dette, noen mindre av det, men de er de samme genene og mange av dem er i samme rekkefølge," ... " Vi trodde de ville bli helt kryptert, men det er de ikke. Det er store deler av det menneskelige genomet som sitter akkurat der i kengurugenomet."
Første dekodede pungdyrgenom avslører 'Junk DNA' overraskelse - 2007
Utdrag: Spesielt fremhever studien de genetiske forskjellene mellom pungdyr som opossum og kenguruer og placenta-pattedyr som mennesker, mus og hunder. ,,,
Forskerne ble overrasket over å finne at pattedyr og pungdyr i stort sett har samme sett med gener for å lage proteiner. I stedet ligger mye av forskjellen i kontrollene som slår gener på og av.

Hvor kunne vi ha lært, bortsett fra Phys.org - 28. september 2014
Utdrag: "Vi har i utgangspunktet de samme 20 000 (30 000?) Proteinkodende gener som en frosk, men likevel er genomet vårt mye mer komplisert, med flere lag av genregulering."
http://www.uncommondescent.com/human-evolution/where-could-we-have-learned-but-from-phys-org/
Likevel er det nøyaktig i disse genetiske reguleringsnettverkene at det finnes størrelsesordener av forskjeller mellom mellom arter:
Evolusjon ved spleising - Sammenligning av genutskrifter fra forskjellige arter avslører overraskende spleisingsmangfold. - Ruth Williams - 20. desember 2012
Utdrag: Et stort spørsmål i evolusjonær biologi hos virveldyr er "hvordan oppstår fysiske og atferdsmessige forskjeller hvis vi har et veldig likt sett med gener som mus, kylling eller frosk?" ...
En ofte diskutert mekanisme var variable nivåer av genuttrykk, men både Blencowe og Chris Burge , fant at genuttrykk er relativt konservert blant arter.
På den annen side viser avisene at de fleste alternative spleisningshendelser er vidt forskjellige mellom selv nært beslektede arter. "De alternative spleisemønstrene er veldig forskjellige, selv mellom mennesker og sjimpanser," sa Blencowe. ,,,
http://www.the-scientist.com/?articles.view%2FarticleNo%2F33782%2Ftitle%2FEvolution-by-Splicing%2F
Genreguleringsforskjeller mellom mennesker, sjimpanser veldig komplekse - 17. oktober 2013
Utdrag: Selv om mennesker og sjimpanser deler .. lignende genomer, har tidligere studier vist at arten utviklet store forskjeller i mRNA (messenger RNA) ekspressjonsnivåer ...
http://www.sciencedaily.com/releases/2013/10/131017144632.htm
"Hvor (sjimpanser og mennesker) virkelig skiller seg ut, og de skiller seg etter størrelsesorden, ligger i genomarkitekturen utenfor proteinkodende regioner. De er veldig, veldig forskjellige. .. Den strukturelle, organisasjonen, de regulatoriske sekvensene, hierarkiet for hvordan ting er organisert og brukt er vidt forskjellige mellom en sjimpanse og et menneske i deres genomer."
Raymond Bohlin (per Richard Sternberg) - 9:29 minutters video: Lenke.
Bilde: Såkalt søppel-DNA
 Om menneskelig opprinnelse: Er genomet vårt fullt av søppel-DNA? Pt 2. - Richard Sternberg PhD. Evolusjonær biologi
Om menneskelig opprinnelse: Er genomet vårt fullt av søppel-DNA? Pt 2. - Richard Sternberg PhD. Evolusjonær biologi
Utdrag: "Her er det interessante når du ser på proteinkodende sekvenser du har i cellen din, det du finner er at de er nesten identiske med proteinkodende sekvenser til en hund, en karpe, en fruktflue , av en nematode. De er praktisk talt de samme og de er utskiftbare. Du kan slå ut et gen som koder for et protein for et indre øreben, for eksempel en mus. Dette er gjort. Og så kan du ta et protein som ligner på det, men fra en fruktflue. Og fruktfluer er ikke virveldyr, og de er absolutt ikke pattedyr. Så de har ikke indre øreben. Og du kan koble det genet inn og gjette hva som skjer? Avkommet til musen vil ha et helt normalt indre øreben. Så du kan bytte ut alle disse filene. Jeg nevner dette for deg fordi når du hører om at vi er 99% like (til sjimpanser), refererer det nesten til de proteinkodende regionene. Når du begynner å lete, og du begynner å sammenligne forskjellige pattedyr. Delfiner, jordvarker, elefanter, sjøfugler, mennesker, sjimpanser, det spiller ingen rolle. Det du finner er at de proteinkodende sekvensene er veldig godt konservert, og det er også mye av DNA som ikke er proteinkodende som også er høyt konservert.
Men når du ser på kromosomene og de båndmønstrene, de strekkodene, (nevnt i begynnelsen av samtalen), er det lik å gå inn i matbutikken. Du ser en haug med svarte og hvite streker, ikke sant? Du har sett en strekkode, og du har sett dem alle. Men disse strekkodene er ikke de samme. .. Her er et eksempel, jordsvin og menneskelige kromosomer. De ser veldig like ut på DNA-nivå når du tar små utdrag av dem. (Likevel) Når du ser på hvordan de er ordnet i et lineært mønster langs kromosomet, viser de seg å være veldig forskjellige (fra hverandre). Så når du kommer til mappen og supermappen og høyere ordrenivå, er det da du finner disse slående forskjellene. Og her er et annet eksempel. De sekvenserer nå kjerne DNA til den atlantiske nese-delfinen. Og da de opprinnelig begynte å sekvensere DNA, var det første de innså at delfin-genomet i utgangspunktet er nesten helt identisk med det menneskelige genomet. Det vil si at det er noen få kromosomomdannelser her og der, du stiller sekvensene opp og de passer veldig bra. Likevel vil ingen hevde, basert på en uttalelse som den, at nese-delfiner er nært knyttet til oss. Vår søsterart hvis du vil. Ingen ville antas å gjøre det. Så du må satse på en annen formodning. Men her er poenget. Du vil se disse utsagnene gjennom litteraturen om hvor vanlige ting er. .. (Delelister er veldig like, men hvordan delene brukes er der du vil finne enorme forskjeller)
Lenke.
Videre, i motsetning til proteinkodende regioner hvor det er en viss 'ikke-katastrofal' toleranse for tilfeldige mutasjoner, viser tilfeldig mutasjoner av genregulerende nettverk alltid å være 'katastrofalt dårlige':
A Listener's Guide to the Meyer-Marshall Debate: Focus on the Origin of Information Question -Casey Luskin - 4. desember 2013
 Utdrag: "Det er alltid en observerbar konsekvens hvis en dGRN (utviklingsgenereguleringsnettverk) -underkrets blir avbrutt. Siden disse konsekvensene alltid er katastrofalt dårlige, er fleksibiliteten minimal, og siden delkretsene alle er sammenkoblet, har hele nettverket den kvaliteten at det bare er én måte for ting å fungere på. Og faktisk utvikler embryoene til hver art bare på en måte. " -
Utdrag: "Det er alltid en observerbar konsekvens hvis en dGRN (utviklingsgenereguleringsnettverk) -underkrets blir avbrutt. Siden disse konsekvensene alltid er katastrofalt dårlige, er fleksibiliteten minimal, og siden delkretsene alle er sammenkoblet, har hele nettverket den kvaliteten at det bare er én måte for ting å fungere på. Og faktisk utvikler embryoene til hver art bare på en måte. " -
Eric Davidson - utviklingsbiolog
http://www.evolutionnews.org/2013/12/a_listeners_gui079811.html
Dermed, der darwinister mest trenger plastisitet i genomet for å være levedyktig som en teori (dvs. utviklingsgenereguleringsnettverk), er stedet der mutasjoner blir funnet å være 'alltid katastrofalt dårlige'. Likevel er det akkurat i dette området av genomet (dvs. regulatoriske nettverk) hvor det er betydelige, 'størrelsesordener', forskjeller mellom til og med antatt nært beslektede arter.
Det er unødvendig å si at dette er det stikk motsatte funnet for hva darwinismen ville ha spådd for det som burde blitt funnet i genomet.
Hvis darwinismen var en normal vitenskap, i stedet for i utgangspunktet å være ateisters ufalsifiserbare 'blinde tro'-religion, burde dette funnet i seg selv ha vært mer enn nok til å forfalske nydarwinistiske påstander.
Av tilleggsnotat til Richard Sternbergs 'strekkoder er ikke det samme' mellom arter sitat. Det viser seg at strekkodemønsteret som Dr. Sternberg antydet er irredusibelt kompleks i sin organisatoriske relasjon til de enkelte gener:
Referert vitenskapelig artikkel om DNA argumenterer for irredusibel kompleksitet - 2. oktober 2013:
Utdrag: Denne artikkelen som ... er en ekte tankegang som viser den irredusible organisatoriske kompleksiteten (forfatterens beskrivelse) av DNA-analog og digital informasjon, at gener ikke er vilkårlig plassert på kromosomet osv. ,,
... For det første er den digitale informasjonen til individuelle gener (semantikk) avhengig av de intergene regionene (som vi vet), som er som analog informasjon (syntaks). Begge typer informasjon er 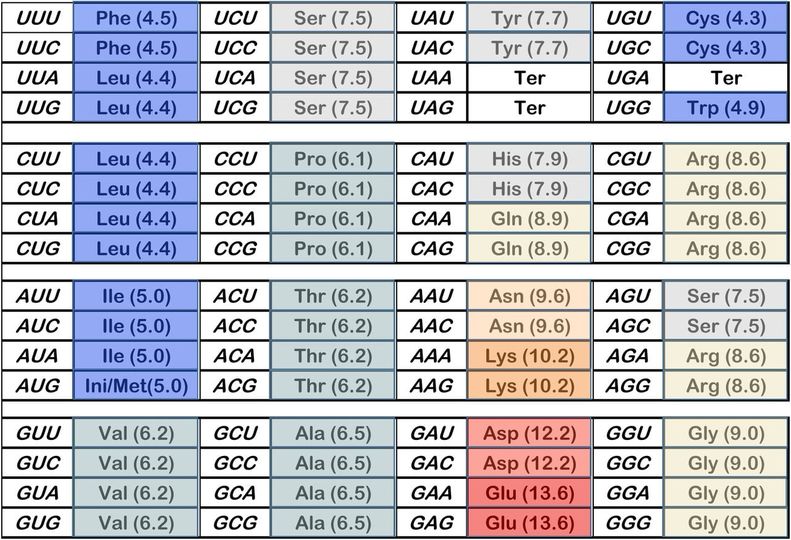 avhengige og selvhenvisende, men du kan ikke få syntaks fra semantikk. Som forfatterne sier, "antar den holistiske tilnærmingen selvreferensialitet (fullstendighet av den inneholdte informasjonen og full konsistens av de forskjellige kodene) som en irreduserbar organisatorisk kompleksitet i det genetiske reguleringssystemet til enhver celle". Kort sagt inneholder den lineære DNA-sekvensen begge typer informasjon. For det andre knytter artikkelen lokal DNA-struktur, til domener, til den generelle kromosomkonfigurasjonen som et dynamisk system som avbryter cellens metabolske signaler. Dette innebærer at posisjonen og organiseringen av gener på kromosomet ikke er vilkårlig .. http://www.christianscientific.org/refereed-scientific-article-on-dna-argues-for-irreducibly-complexity/
avhengige og selvhenvisende, men du kan ikke få syntaks fra semantikk. Som forfatterne sier, "antar den holistiske tilnærmingen selvreferensialitet (fullstendighet av den inneholdte informasjonen og full konsistens av de forskjellige kodene) som en irreduserbar organisatorisk kompleksitet i det genetiske reguleringssystemet til enhver celle". Kort sagt inneholder den lineære DNA-sekvensen begge typer informasjon. For det andre knytter artikkelen lokal DNA-struktur, til domener, til den generelle kromosomkonfigurasjonen som et dynamisk system som avbryter cellens metabolske signaler. Dette innebærer at posisjonen og organiseringen av gener på kromosomet ikke er vilkårlig .. http://www.christianscientific.org/refereed-scientific-article-on-dna-argues-for-irreducibly-complexity/
Dette har vært et ganske langt innlegg, (til og med for meg), men forhåpentligvis for den fordomsfrie personen som ærlig talt prøver å se om enten ID eller darwinisme er sant, har dette innlegget gjort det klart at ny-darwinistiske forklaringer er sterkt mangelfulle på flere forskjellige nivåer for å forklare den fantastiske integrerte kompleksiteten vi ser i livet, og at ID-forklaringer langt er den mest tilfredsstillende forklaringen på den fantastiske integrerte kompleksiteten vi ser.
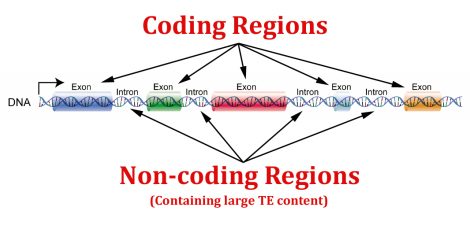
Duoner: Parallell genkode trosser evolusjon 1
Forskere har nettopp karakterisert en ny, tidligere skjult genetisk kode innebygd i de samme seksjonene av gener som koder for proteiner - som fullstendig trosser alle naturalistiske forklaringer på deres eksistens. (1)
I tillegg til å levere mange forskjellige typer genetisk kode som regulerer funksjon, gir genomet også svært komplekse kodede maler for å lage et bredt mangfold av funksjonelle RNA-molekyler og proteiner.
Proteinkodende gener - de som inneholder nøkkelinformasjonen for å lage proteiner - inneholder den mest studerte typen genetisk kode. Noen av de viktigste kodene i gener er eksonene, som spesifiserer den faktiske malen for proteinsekvenser.
I eksoner danner tre påfølgende DNA-bokstaver det som kalles et kodon, og hvert kodon tilsvarer en spesifikk aminosyre i et protein. Lange sett med kodoner i gener inneholder den proteindannende informasjonen som ender med å bli oversatt til hele proteiner som kan ha hundrevis av aminosyrer.
 Før denne studien var forskere klar over at proteinkodende regioner i gener hadde mystiske signaler annet enn kodoner som fortalte cellemaskineriet hvordan man skulle regulere og behandle RNA-transkripsjoner (kopier av gener) før de produserte proteinet. Forskere trodde opprinnelig at disse regulatoriske kodene og proteinmalkodene som inneholder kodonene, fungerte uavhengig av hverandre.
Før denne studien var forskere klar over at proteinkodende regioner i gener hadde mystiske signaler annet enn kodoner som fortalte cellemaskineriet hvordan man skulle regulere og behandle RNA-transkripsjoner (kopier av gener) før de produserte proteinet. Forskere trodde opprinnelig at disse regulatoriske kodene og proteinmalkodene som inneholder kodonene, fungerte uavhengig av hverandre.
I virkeligheten viste de nye resultatene at disse kodene faktisk fungerer hver for seg og sammen. Mens ett sett med kodoner spesifiserer rekkefølgen av aminosyrer for et protein, spesifiserer den samme sekvensen av DNA-bokstaver også hvor det er nødvendig for cellulært maskineri (transkripsjonsfaktorer) å binde seg til genet for å lage RNA-transkripsjonen som koder for et protein. Som et resultat av denne nye oppdagelsen har disse dual-funksjon kode stedene i eksoner blitt kalt 'duoner'. Forskere rapporterte bare i fjor at transkripsjonsfaktorer festet seg på noen eksoner i gener, men forsto ikke dette dobbeltkodesystemet før nå. (2)
Menneskesinnet sliter med å forstå den generelle kompleksiteten til den genetiske koden - spesielt bevisene som kommer fram som viser at noen gener har seksjoner som kan leses både forlengs og baklengs. (3) Noen gener overlapper deler av andre gener i genomet, og nå er det blitt avslørt at mange gener har områder som inneholder doble koder i samme sekvens. (1,4)
Selv de mest avanserte programmerere kan ikke komme i nærheten av å matche den genetiske kodens utrolige informasjonstetthet og forvirrende kompleksitet. En allmektig Skaper ser ut til å være den eneste forklaringen på denne forbløffende mengden tilsynelatende uendelig bioteknologi i genomet. {Det er en slutning grunnet på menneskelige erfaring, at all spesifisert kompleksitet viser seg å stamme fra et intelligent opphav -oversetters tilføyelse.}
Utfyllende notater:
I motsetning til hva mange tror, støtter ikke fossilopptegnelsen i sin helhet, hypotesen om felles avstamning (Ulikhet går konsekvent foran mangfold i fossilregisteret)
misforhold
1. mangel på likhet eller likhet; ulikhet; forskjell
Faktisk gir 'argumentet fra form' oss også veldig godt bevis på at vi hver må ha en sjel for å forklare hvordan milliard og billioner av proteinmolekyler i en menneskekropp muligens kan sammenføye seg som en samlet helhet 'nøyaktig hele livet' , og ikke et øyeblikk lenger (Talbott).
Kroppsplaner er, i motsetning til ny-darwinistiske forutsetninger, ganske enkelt ikke reduserbare til DNA (punktum) Dette funnet gjengir nesten ethvert Darwinistisk argument for felles forfedre, basert på DNA alene, som omstridt og tomt.
A Big Problem for Common Descent: Hundreds of "Active 'Foreign' Genes" Don't Fit the Standard Evolutionary Phylogeny
http://www.evolutionnews.org/2015/03/a_big_problem_f094701.html
Some Problems in Proving the Existence of the Universal Common Ancestor of Life on Earth
http://www.hindawi.com/journals/tswj/2012/479824/
A Primer on the Tree of Life
http://www.ideacenter.org/contentmgr/showdetails.php/id/1481
Multiple Overlapping Genetic Codes Profoundly Reduce the Probability of Beneficial Mutation George Montañez 1, Robert J. Marks II 2, Jorge Fernandez 3 and John C. Sanford 4 - May 2013 George Montañez 1, Robert J. Marks II 2, Jorge Fernandez 3 and John C. Sanford 4 - May 2013
Utdrag: Det er nesten allment anerkjent at gunstige mutasjoner er sjeldne sammenlignet med skadelige mutasjoner [1-10] .. Det ser ut til at gunstige mutasjoner kan være for sjeldne til å faktisk tillate en nøyaktig måling av hvor sjeldne de er [11].
Referanser:
Kontra: Og dette er blitt til ved tilfeldighet

1. Kibota T, Lynch M (1996) Estimate of the genomic mutation rate deleterious to overall fitness in E. coli . Nature 381:694-696.
2. Charlesworth B, Charlesworth D (1998) Some evolutionary consequences of deleterious mutations. Genetica 103: 3-19.
3. Elena S, et al (1998) Distribution of fitness effects caused by random insertion mutations in Escherichia coli. Genetica 102/103: 349-358.
4. Gerrish P, Lenski R N (1998) The fate of competing beneficial mutations in an asexual population. Genetica 102/103:127-144.
5. Crow J (2000) The origins, patterns, and implications of human spontaneous mutation. Nature Reviews 1:40-47.
6. Bataillon T (2000) Estimation of spontaneous genome-wide mutation rate parameters: whither beneficial mutations? Heredity 84:497-501.
7. Imhof M, Schlotterer C (2001) Fitness effects of advantageous mutations in evolving Escherichia coli populations. Proc Natl Acad Sci USA 98:1113-1117.
8. Orr H (2003) The distribution of fitness effects among beneficial mutations. Genetics 163: 1519-1526.
9. Keightley P, Lynch M (2003) Toward a realistic model of mutations affecting fitness. Evolution 57:683-685.
10. Barrett R, et al (2006) The distribution of beneficial mutation effects under strong selection. Genetics 174:2071-2079.
11. Bataillon T (2000) Estimation of spontaneous genome-wide mutation rate parameters: whither beneficial mutations? Heredity 84:497-501.
http://www.worldscientific.com/doi/pdf/10.1142/9789814508728_0006
Oversettelse og bilder ved Asbjørn E. Lund